ENaC - An Overview
Structural biology is ultimately the study of particular domains and residues, but this knowledge most interesting and useful in the context of physiological processes. As such, it can be difficult to order the chapters of an introduction. Placing the structure before the function grounds discussion of mutations and disease, but leaves the structure unmoored from biomedical relevance. Conversely, placing function first leaves the reader without a clear understanding of which domains are important for which reasons, which is vital for a focused discussion of the protein.
To dodge the question of which is the cart and which is the course, I present here an exceedingly brief (and less heavily cited) discussion of ENaC structure and function. Everything covered here is covered in more detail in the Introduction. ENaC biology is discussed in Chapter 1, while details on the ENaC/DEG family and ENaC structure can be found in Chapter 2.
A bit of ion channel terminology
Ion channels are essential components of cellular life, present in everything from bacteria on up to humans. The minimal ion channel spans the lipid bilayer and allows ions to flow through a pore down their concentration gradient. We can measure the flow of ions through a single channel as a current (called the unitary current).
However, most ion channels have some mechanism by which the flow of ions can be controlled. We thus say an ion channel can be “open” or “closed”, i.e., it can allow ions to flow through itself or not. This process is called gating, and is most often controlled by either voltage (voltage-gated ion channels) or some ligand (ligand-gated ion channels).
We can further quantify a channel’s “openness” by how much time it spends in the open state vs. the closed state. A channel with an open probability (PO) of 0.75 spends approximately 75% of its time in the open state and 25% of its time in the closed state.
Open probability and unitary currents are both properties of single channels. Groups of channels (for instance, all of the channels in a cell) do not have a measurable PO, since the group will not be homogeneous.
For instance, say there is a collection of 100 ion channels. Twenty of those channels are bound to ligand, and seventeen of those channels are open. Eighty channels are not bound to ligand, and twenty-two of those channels are open. Do we only consider the ligand-bound channels, making the PO \(\frac{17}{20}\)? Or maybe we should consider only the apo channels, making it \(\frac{22}{80}\)? Or perhaps we consider the whole population: \(\frac{39}{100}\)? Or is it maybe the mean PO of both groups, \(\frac{\frac{17}{20} + \frac{22}{80}}{2}\)?
In practice, we’d probably describe this set of 100 channels as two populations: the open population with a PO of 0.85 and the closed population with a PO of 0.275. Ligand binding would move a channel from the closed to the open population, and ligand dissociation would move it back to the closed population.
Typical electrophysiology experiments measure the macroscopic current: how many ions flow through all of the channels of a given type in a cell. This is represented as I, with the current due to a particular ion, e.g., sodium as INa+. Ion channels are often selective for a particular ion or set of ions. For instance, a sodium-specific ion channel will allow Na+ but not other ions through, while a cation-selective channel will allow Ca2+ and K+ but not Cl-. Macroscopic current is affected by the number of channels in each gating state, the PO of those states, the unitary current of the open channels, and the number of channels in the membrane (N). Further review is available from Hille or Ackerman and Clapham1,2.
The ENaC/DEG family
The epithelial sodium channel (ENaC) is, perhaps obviously, a member of the ENaC/DEG family of ion channels. These channels are trimeric, pass cationic currents, are voltage-insensitive, and are inhibited by the small molecule amiloride (and its derivatives). Amiloride blocks current through ENaC/DEG channels by binding a surface in the extracellular to the selectivity filter and blocking the pore.
The overall topology of these channels is similar, with the greatest sequence (and, presumably, structural) diversity found in the distal ECD (furthest from the membrane). Current structural study suggests a conserved overall gating mechanism as well.
Aside from ENaC, the best-studied members of the ENaC/DEG family are the eponymous degenerin (DEG) channel and the acid-sensing ion channel, ASIC. DEG is expressed in C. elegans and is involved in mechanosensation. ASIC is expressed in a variety of organisms (including humans) and opens in response to acidic extracellular conditions. ASIC has by far the best-described structure of the family.
ENaC Structure
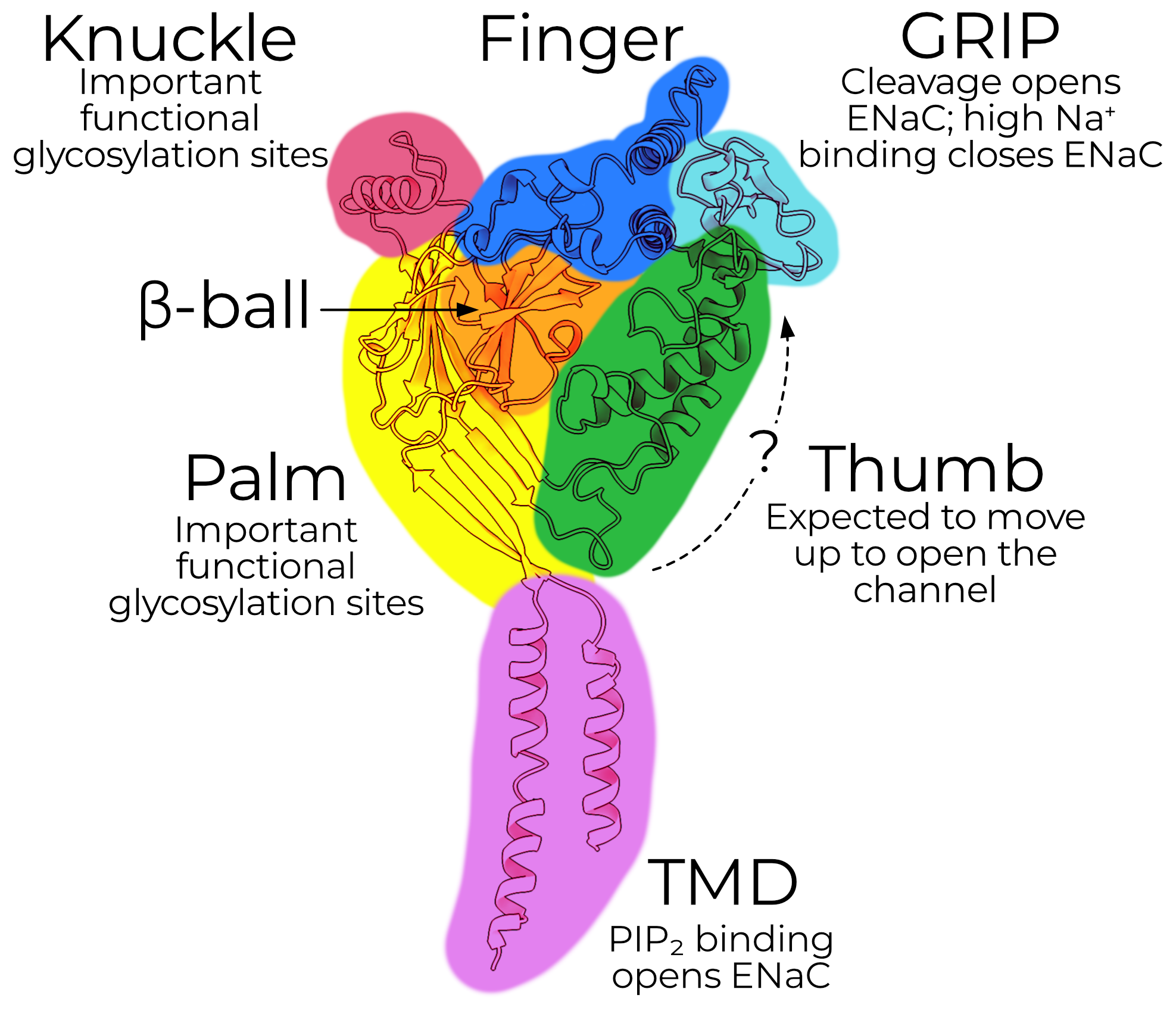
ENaC is a triheteromeric ion channel comprising three subunits, the α, β, and γ subunits. Each subunit shares an overall similar topology, and so I describe them here without reference to a specific one.
The transmembrane domain, or TMD, contains the pore and selectivity filter. The extracellular domain, or ECD, contains domains important for regulating open probability (PO) and other functional characteristics. There are also short intracellular N- and C-terminal tails which regulate surface expression.
The ECD is further subdivided into six domains, named after the parts of a hand. The palm and β-ball form a rigid backbone and make the plurality of the contacts with the other subunits. The knuckle, finger, and thumb likely move during channel gating and contain important glycosylation sites.
The GRIP domain is unique to ENaC. ENaC opens when the GRIP domain is excised from the channel. The canonical proteases responsible for channel opening are: furin, which cuts the α subunit twice and γ subunit once; and prostasin, which cuts the γ subunit again.
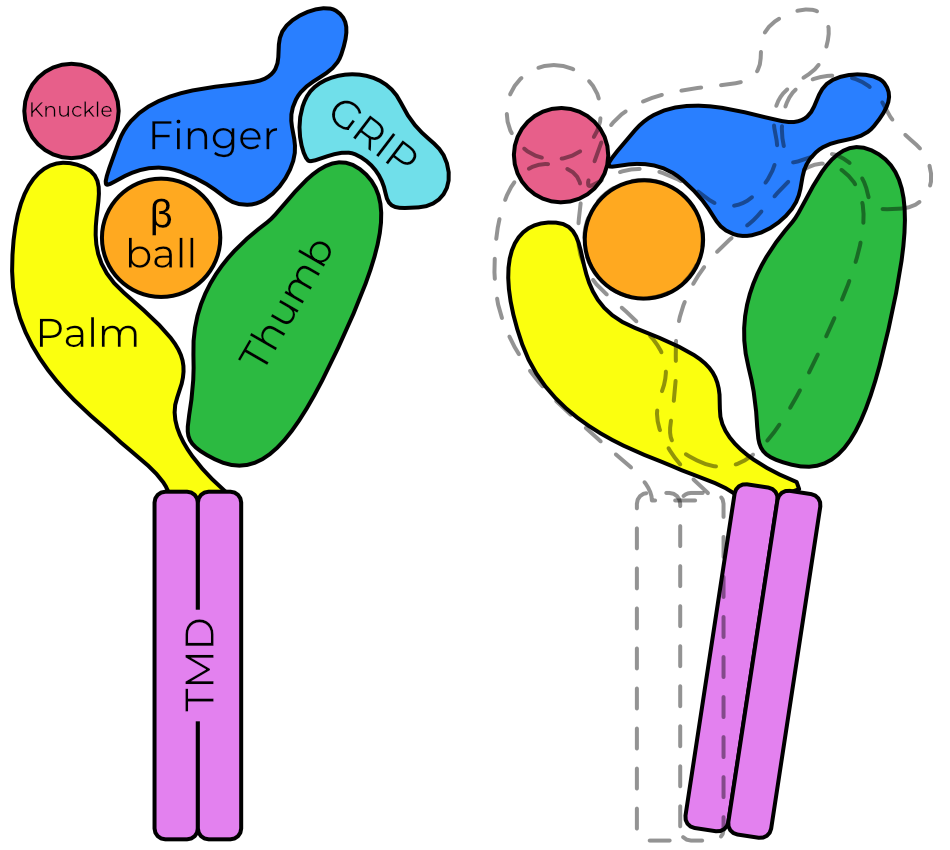
Removal of the GRIP domain is thought to allow the finger and thumb to collapse together. This movement is coordinated with movement of the palm away from the pore axis, which in turn opens a fenestration just above the membrane. Ions enter at this fenestration and pass down a pore formed by the second transmembrane helix, TM2.
Along this pore, ions larger than sodium are filtered out by size. It is thought that this selectivity filter is formed by a three-residue loop connecting swapped domains of TM2, but an un-swapped conformation is also possible.
The above summary of ENaC’s presumed gating mechanism is based largely off cross-linking electrophysiology studies and ASIC’s well-described gating mechanism. In ASIC, protons allow for finger/thumb collapse (recall that ASIC does not have a GRIP domain). However, ASIC rapidly desensitizes and ENaC does not.
ENaC Function and Regulation
The regulation and function of ENaC in the body is not entirely understood, but broadly, ENaC functions as the rate-limiting step of sodium reabsorption in tissues which express it. Thus, control of ENaC expression also controls water retention and reabsorption in those tissues.
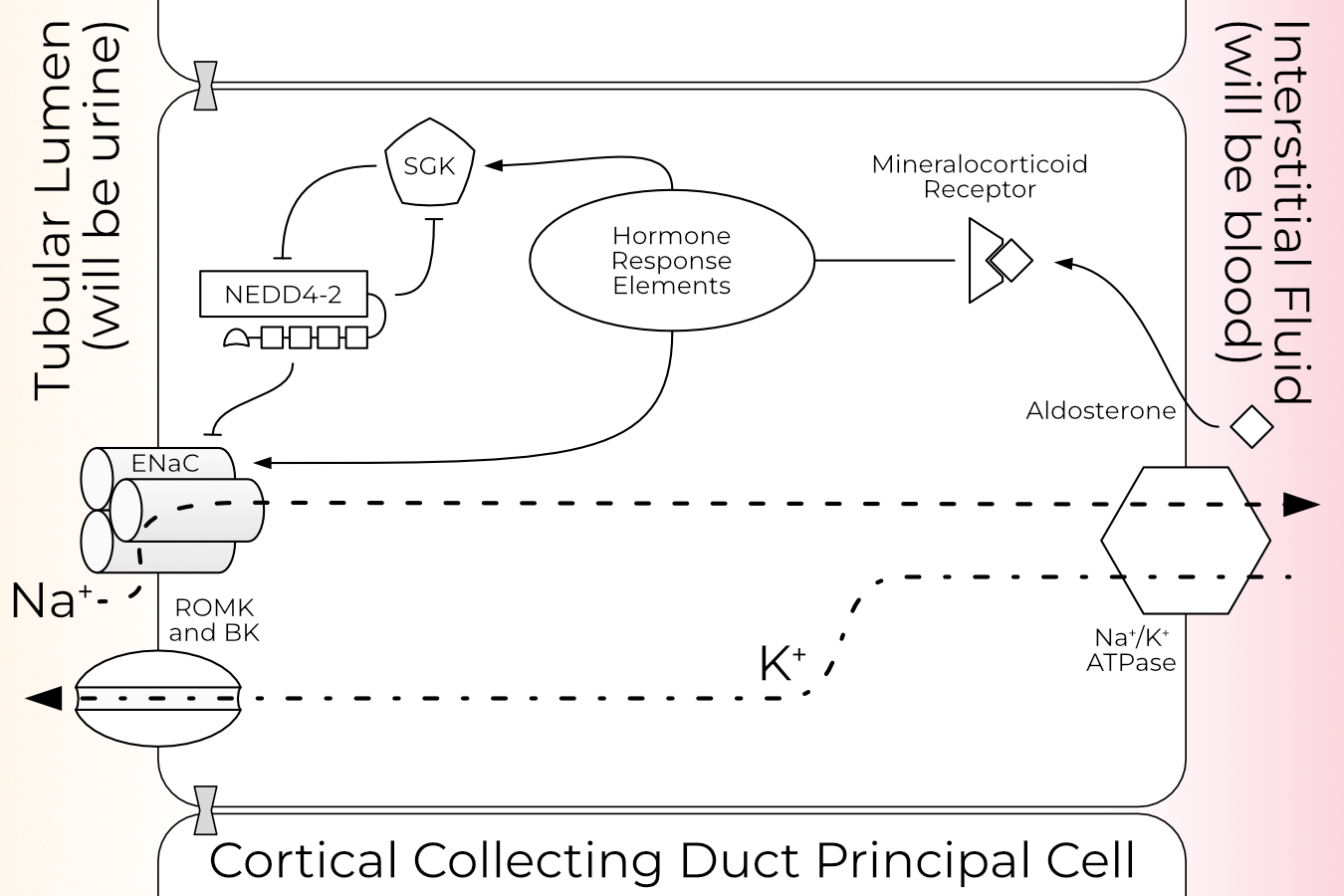
ENaC is best studied in the kidney, where it plays a pivotal role in blood pressure regulation. When the kidney detects reduced blood pressure, it initiates a cascade which results in increased circulating levels of the mineralocorticoid hormone aldosterone. Aldosterone increases overall expression of ENaC, the proportion of ENaC at the cell surface, and the proportion of channels which are open (i.e., have an excised GRIP domain). For more detail on aldosterone and ENaC regulation in the kidney, see Section 1.2.1.

Cells with more ENaC at the surface allow more sodium to pass from the lumen to the interstitial fluid. This in turn results in increased water retention, thus raising blood volume and pressure.
Since the channel is activated primarily by proteolysis, ENaCs cannot close again once opened. Open channels are tagged for retrieval from the plasma membrane and degradation via ubiquitination on the N-termini each of the three subunits. Additionally, ENaC is sensitive to membrane composition and requires PIP2 to open.
This is, I believe, enough of a background to move on to a full introduction of the topics covered in the remaining document. The reader is also encouraged to check the glossary for unfamiliar terms and abbreviations.