2 ENaC Currents, Structure, and Gating
In this chapter I begin with an overview of ENaC electrophysiology. From there, I move into a discussion of current knowledge about ENaC’s structure/function relationships. Finally, I provide a brief overview of proteins which directly modify ENaC’s electrophysiological properties (rather than expression levels, which are covered in Section 1.2).
2.1 ASIC
ENaC is a member of the ENaC/DEG superfamily of sodium-selective ion channels. These ion channels share a common ancestor as far back as Cnidaria and, as their long evolutionary history might imply, fill a wide variety of roles in the organisms that express them1. The structures of ENaC/DEG family members solved so far also hold to a common topology: the ion channels are trimeric, with large extracellular domains, two transmembrane helices per subunit, and short intracellular tails.
The member of this family of which we have the greatest structural understanding is the acid-sensing ion channel (ASIC). All of the gating states of this channel have been solved, and a great deal is known about its gating mechanism and the conformational changes associated with channel opening, desensitization, and closing. Since existing functional study of ENaC suggests similar conformational changes, most researchers assume that the conformational changes associated with ENaC’s gating mechanism will be similar. In this section I provide the reader with a brief introduction to ASIC and important similarities and differences between it and ENaC.
The ASICs are themselves a family of ion channels, and the properties of each channel differ slightly from the others. There is also evidence that different ASIC proteins can form heterotrimeric ASIC channels. However, the only ASIC for which we have structural information is ASIC1a, and so I refer to that channel as ASIC for the rest of this document. I will explicitly refer to the ASIC family as necessary.
2.1.1 Gating
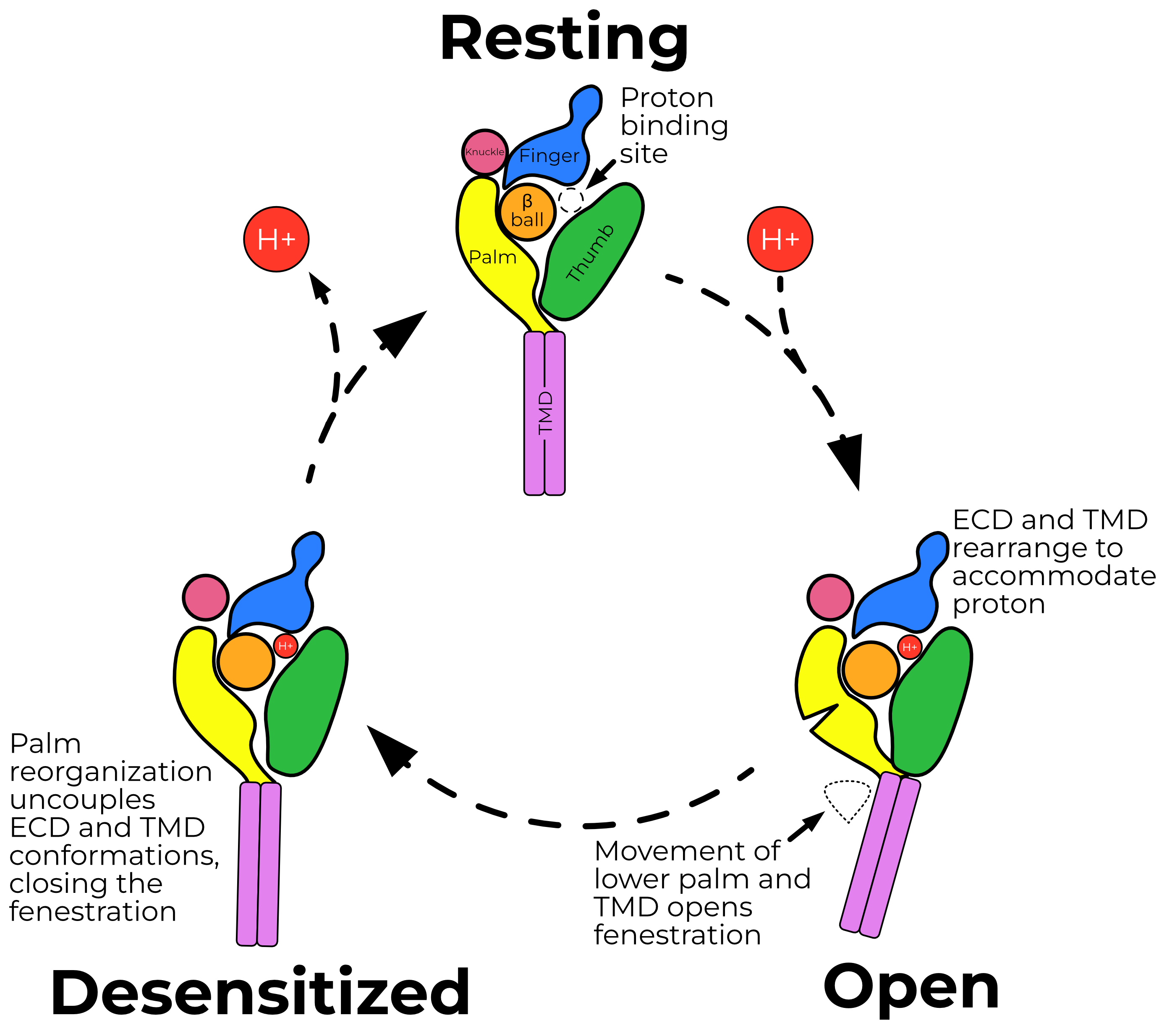
ASIC’s gating cycle involves three states: resting, open, and desensitized (Figure 2.1). ASIC opens when its extracellular domain is exposed to acidic solution, with a half-maximum activation at pH 5.82. This peak current then desensitizes with a time constant of 0.64 — a marked difference from ENaC currents, which do not desensitize.
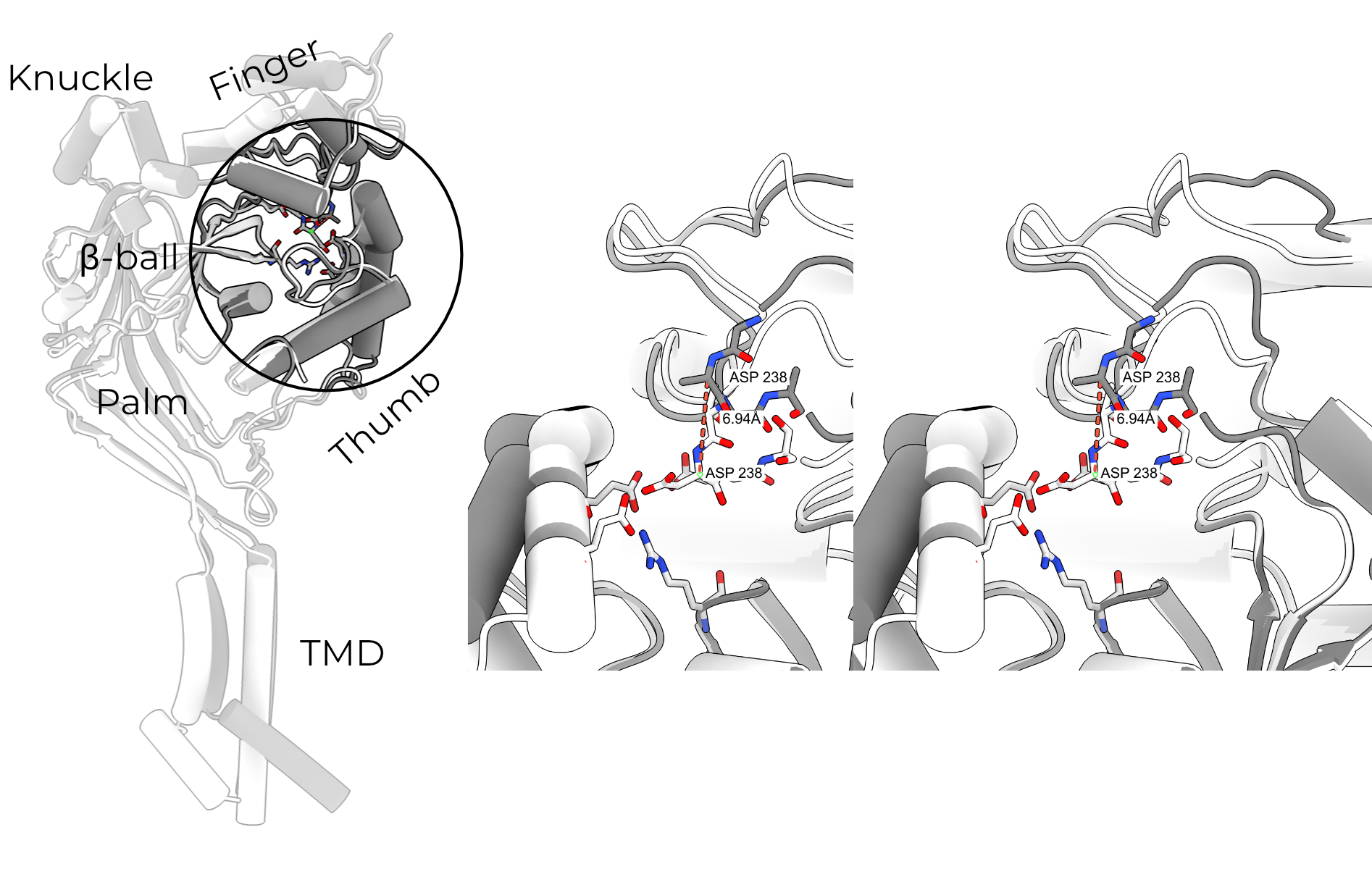
Protons open the channel by binding an acidic pocket located between the finger, thumb, and β-ball domains (Figure 2.2). Binding of protons to titratable residues in this pocket result in the collapse of the finger and thumb. This collapse is in turn propagated through the thumb and palm to the TMD, which rotates and translates about the pore axis (Figure 2.3). This movement opens a fenestration just above the cell membrane, through which ions enter the pore (Figure 2.4).

The pore is lined by the second transmembrane helix from each subunit. Approximately two-thirds of the way down the pore, the helix breaks and short loop connects a domain swap to the adjacent TM2 helix (Figure 2.5). This loop, called the GAS belt due to its one-letter code sequence, forms the selectivity filter of ASIC.
Unlike ENaC, ASIC desensitizes. Rapidly upon opening, a short loop in ASIC’s palm flips to a desensitized conformation, which decouples the ECD and TMD. Thus, the ECD remains proton-bound and open, while the TMD returns to a closed conformation and obstructs the fenestration, preventing ions from passing through.
2.2 ENaC Electrophysiology
ENaC currents are characterized by high Na+/K+ selectivity, high affinity for amiloride, and a desensitization-like process called sodium self-inhibition (SSI). Here I cover each of these facets and connect them to the following discussion of ENaC structure.
2.2.1 The pore and selectivity filter
ENaC is highly selective for Na+ over K+, with an estimated INa+/IK+ well over 1003,4. The precise number depends on the study, since potassium currents through ENaC are often difficult to detect at all. ENaC does not pass appreciable H+ currents until the extracellular solution reaches a pH of 5 or lower5.
Ions enter the channel through a fenestration just outside the membrane and travel down a pore formed by the second transmembrane helices of all three subunits, just as in ASIC4,6. As discussed further in Section 2.3.2.3, ENaC’s selectivity filter is composed of a triplet of residues in a domain-swapping loop between adjacent transmembrane helices. Based on structures of related channels, this domain swap is expected to occur approximately halfway through the membrane7. The third residue of the α loop (a serine) plays an outsize role in determining channel selectivity8.
ENaC achieves high specificity for sodium by “sieving” ions, physically preventing larger cations from moving down the pore. The selectivity filter is so small that Na+ ions must be dehydrated before passing through8. Because ENaC separates by size, Li+ currents are slightly larger than Na+ currents (ILi+/INa+ ≅ 1.5). The energetic cost of this sieving results in a low single-channel conductance (approximately 5 pS)3.
2.2.2 Amiloride and other inhibitors
The ENaC/DEG family of ion channels are all inhibited (to varying degrees) by the small molecule amiloride. Amiloride has the highest affinity for ENaC, with an IC50 of about 100 nM. More hydrophobic derivatives (e.g., benzamil or phenamil) bind approximately ten times tighter, indicating that amiloride binds ENaC at a hydrophobic patch (amiloride derivatives are reviewed well by Kleyman and Cragoe9).
Amiloride binds just above (i.e., extracellular to) the selectivity filter in a voltage-dependent manner4,10. Some mutagenesis studies indicate that amiloride interacts primarily with the β and γ subunits, while others (and evolutionary analysis) are more consistent with an extended interaction surface including the α subunit4,6,11. Studies with sulfhydryl reagents suggest that the binding surface is only accessible when the channel is in the open state6.
2.2.3 PO and sodium self-inhibition
ENaCs are characterized by long dwell times in both the open and closed states and a low likelihood of switching states3. Proteolytic cleavage of the extracellular domain of the channel pushes the channel from a state with a PO of nearly zero to a constitutively open state12. Conversely, intracellular acidification and Ca2+ both reduce channel PO13,14. Unlike the related Acid Sensing Ion Channel (ASIC), ENaC is not directly opened by changes in extracellular pH.
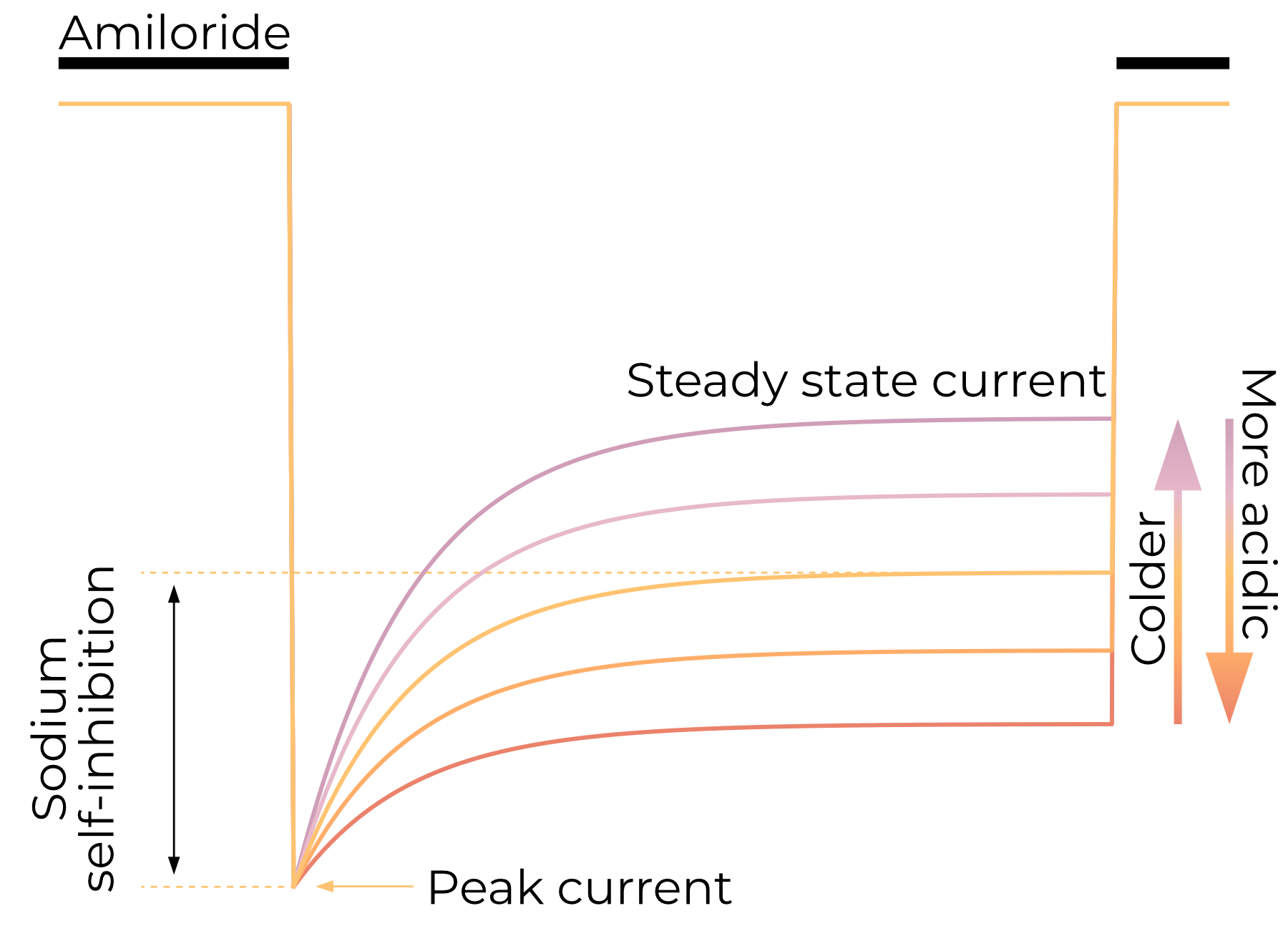
As they do not have a ligand per se, ENaCs do not desensitize in the traditional sense. Steady-state macroscopic currents are, however, lower than peak currents15. This reduction in sodium current on the scale of seconds is called sodium self-inhibition (SSI) and is an important characteristic of ENaC electrophysiology.
SSI reduces ENaC currents as a direct result of extracellular Na+ concentrations, not the currents themselves16. Lower temperatures reduce the magnitude of SSI, resulting in greater steady-state ENaC currents16,17. ENaC is believed to have only a single open and single closed state3,18.
As discussed in Section 2.4.1, ENaC is activated by proteolytic removal of a portion of its extracellular domain called the Gating Relief of Inhibition by Proteolysis (GRIP) domain. This increase in current is a result of SSI relief16,19. It is reasonable to expect, then, that the sodium-binding site is composed at least in part by the GRIP domain. Indeed, an acidic pocket fits this description and binds as-yet-unidentified ions20,21. While extracellular pH does not open ENaC, more acidic extracellular solutions reduce SSI magnitude, resulting in a larger steady-state current22. More basic extracellular solutions result in greater SSI magnitude and therefore lower steady-state currents.
Surprisingly, despite its effect on SSI magnitude, pH does not influence Na+ Ki. If solvent-exposed acidic residues do participate in sodium-binding for SSI, one would expect their protonation to reduce the affinity for sodium. Additionally, the effect of pH on SSI magnitude seems to be species-dependent, further complicating the role of the acidic pocket.
2.3 ENaC Structure
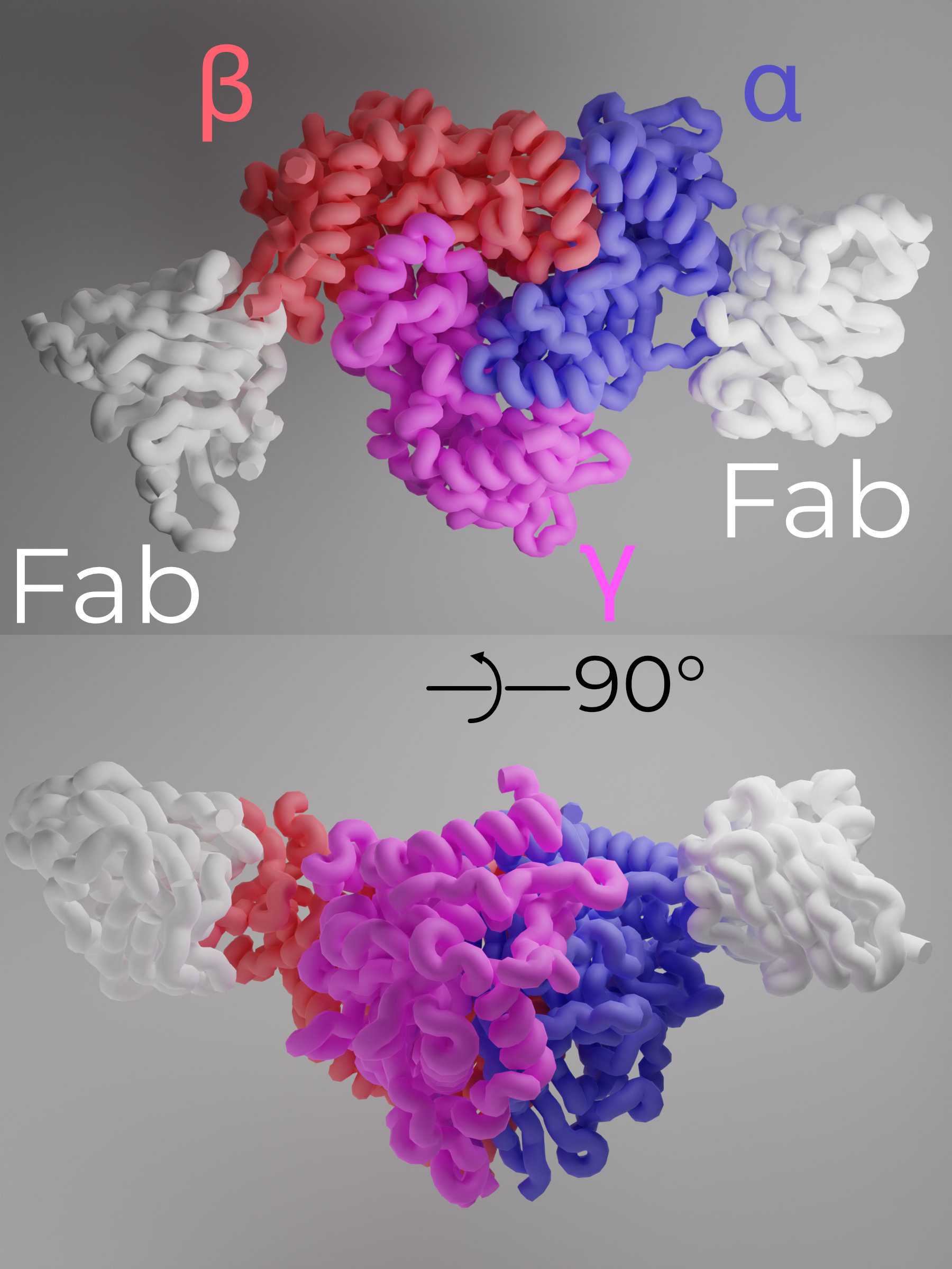
ENaC is a heterotrimeric protein comprising one copy each of αENaC, βENaC, and γENaC arranged counterclockwise when viewed from the extracellular side (Figure 2.7)23,24. Each subunit is approximately 30% identical to the others, and all three share a topology common to the entire ENaC/DEG family. In this section, I will cover structure/function relationships for the two main sections of ENaC: the extracellular and transmembrane domains (ECD and TMD, respectively).
I will also make extensive analogy to a closely related channel, the acid-sensing ion channel (ASIC). Throughout this document, when I refer to ASIC, I specifically mean ASIC1 unless I specify a different paralog. ASIC is another member of the ENaC/DEG family which is activated by protons. I recommend Yoder and colleagues’ excellent overview of ASIC’s gating mechanism to interested readers25.
The first structures of a third ENaC/DEG family member, FMRFamide-gated sodium channel 1 (FaNaC 1; hereafter FaNaC), were published during the production of this document26. This is exciting for two reasons. First, FaNaC is much more closely related to ENaC than is ASIC1. Second, FaNaC shares its overall architecture with ENaC and ASIC, aside from divergence at the ligand-recognition region of the distal finger. These similarities further support a shared overall gating mechanism among the ENaC/DEG family.
2.3.1 Stoichiometry
To date, all solved structures of the ENaC/DEG family are trimeric. Prior to the first structure of ASIC, functional studies of the channel suggested a tetrameric, ααβγ channel — Kashlan and Kleyman discuss the abandonment of the tetrameric hypothesis well27. Although homomeric αENaC channels are competent to pass very small amiloride-sensitive currents, expressing all three subunits results in currents over 100 times larger than the α-only channels23. Thus, the canonical ENaC channel is the αβγ heterotrimer.
As discussed in Section 1.1, it is not clear that this canonical channel is the only one expressed in the body. No tissue assayed thus far expresses all three subunits in equal amounts (in some cases, one or more subunits are not expressed at all). Aldosterone rarely induces expression of all three channels equally. More strikingly, the regulatory protein SPLUNC1 binds to βENaC in the lumen and induces internalization of α- and γ- (but not β-) ENaC28. It is not clear how a small (26 kDa), secreted protein could separate βENaC from the α and γ subunits on its own. Indeed, the same study indicates that βENaC becomes less mobile upon SPLUNC1 binding, suggesting coupling to other, larger proteins. A deeper investigation of the various chaperones and processes involved in ENaC assembly and dissociation would be interesting but, unfortunately, there is only a small body of existing literature on the topic. For our purposes, ENaC is the αβγ heterotrimer.
2.3.2 Functional Domains
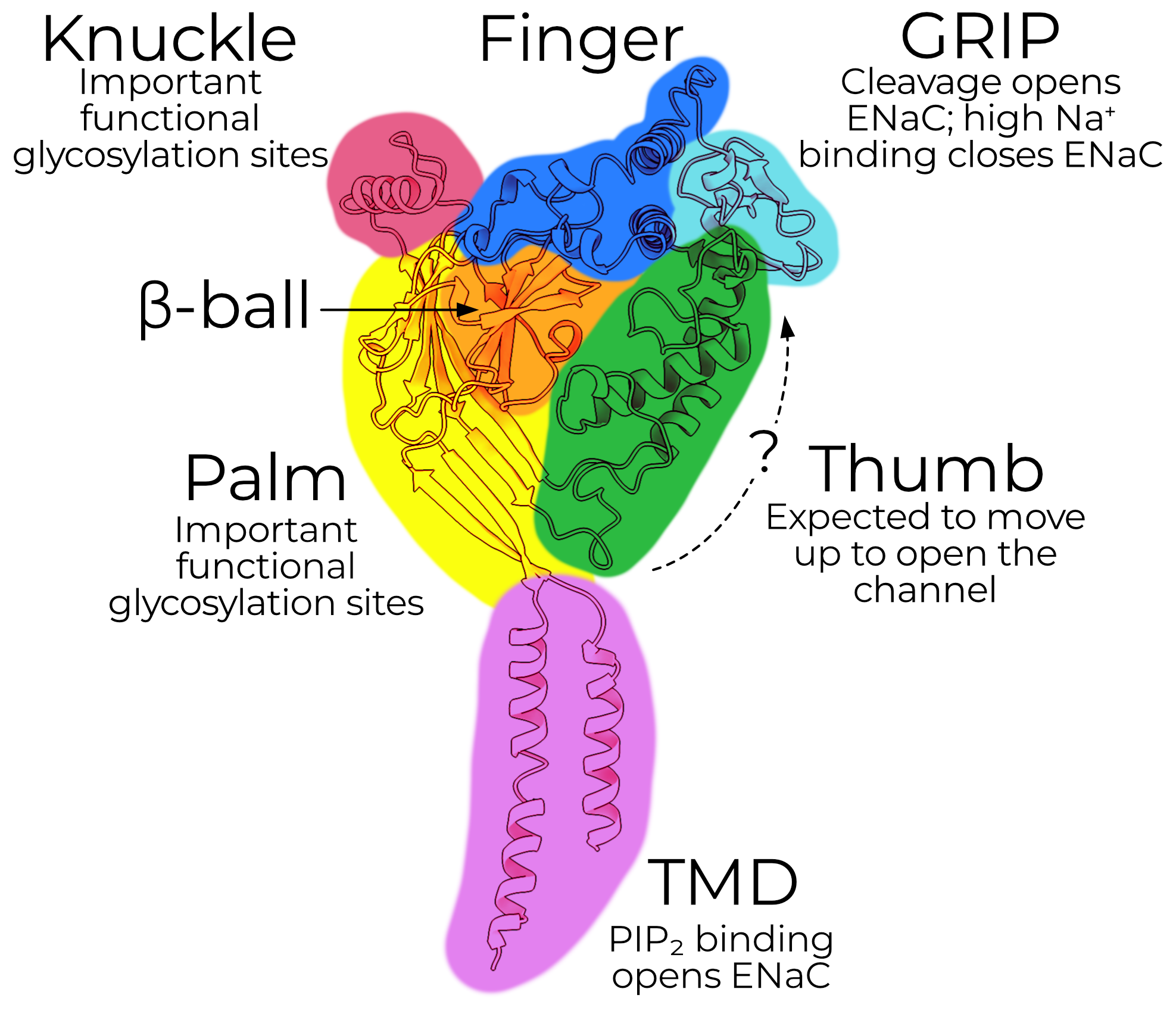
Each subunit has a mass of approximately 75 kDa, with short intracellular N- and C-terminal tails and two transmembrane helices (TM1 and TM2, which together form the transmembrane domain, TMD). The extracellular domain (ECD) comprises the bulk of the protein (approximately 50 kDa), which is further subdivided into domains named for their vague resemblance to a left hand grasping a ball (Figure 2.8). Uncleaved channels typically run a bit higher (between 90 and 120 kDa) on SDS-PAGE due to SDS binding and extensive glycosylation (Section 2.3.3).
2.3.2.1 Scaffolding — the palm, β-ball, and knuckle
The palm (yellow) comprises a twisted β-sheet of seven β-strands (β1, 3, 6, and 9–12). The β-ball forms (as one might expect) a tightly twisted ball of four β-strands: β2, 4, 5, and 7. The knuckle, comprising only a single α-helix (α6) is connected to the extracellular end of the palm.
Although the palm does lie along the pseudo-threefold symmetry axis, the ion permeation pathway of ASIC suggests that it is probably not involved in passing Na+ currents29. The palm instead forms the structural scaffold of each subunit, connecting the finger and knuckle at the extracellular edge of the domain to the thumb at the membrane-adjacent end. About a third of each subunit’s buried surface area (1,667 out of 4,662 Å2 for αENaC) is in the palm, despite it containing only about a fifth of the modeled residues. Given this outsize role in establishing channel topology, it is no surprise that the palm is highly conserved among the ENaC/DEG family and plays a pivotal role in channel gating.
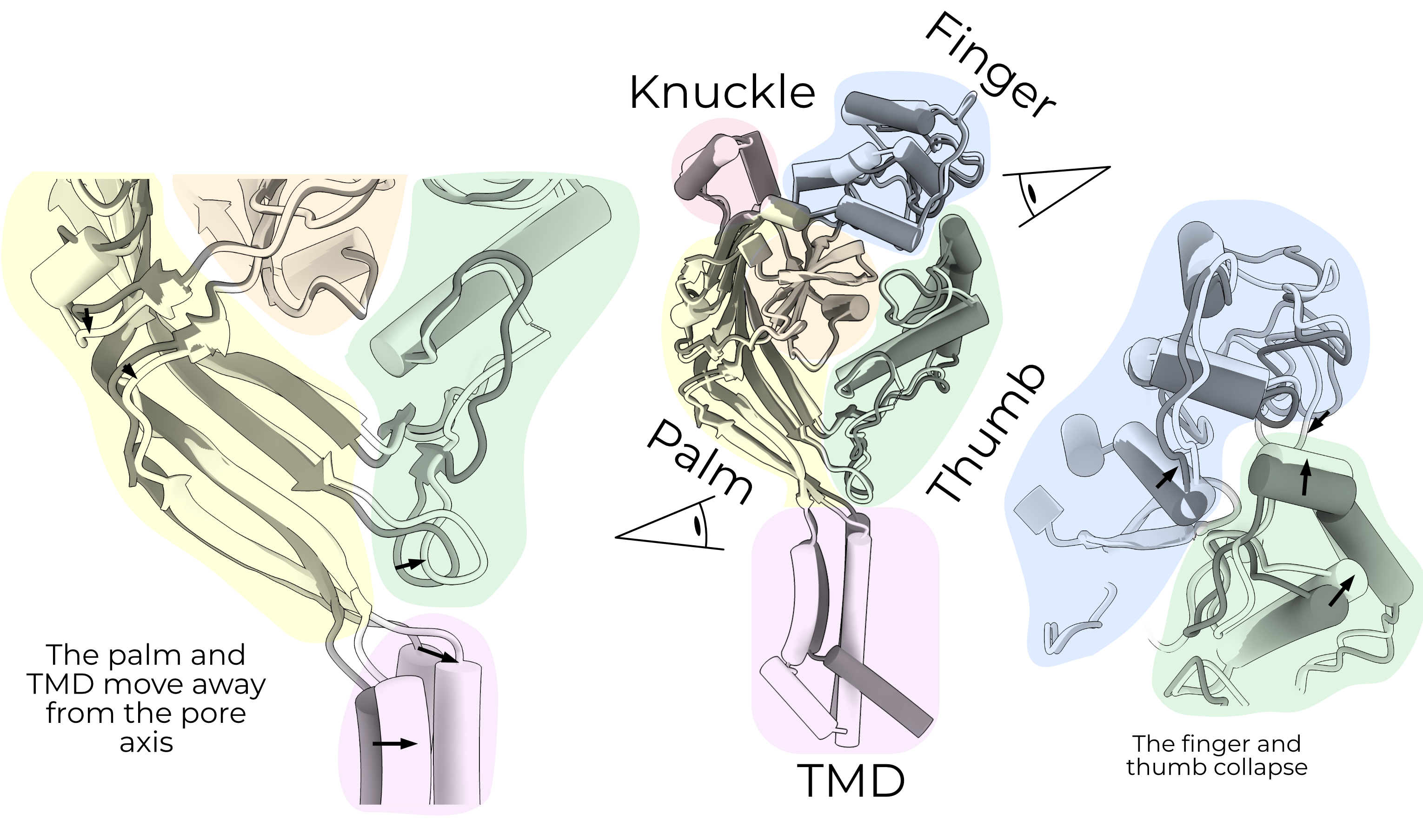
When ASIC opens, the palm pivots around the β-ball (orange) and moves inward by approximately 4 Å7. This movement is coupled to the TMD, opening the pore. The same overall mechanism is expected in ENaC and has support from cross-linking studies30. The α and β palm domains and the α knuckle also host glycosylation sites which are required for mechanosensitive gating (Section 2.4.3). Unfortunately, the research on ENaC glycosylation is relatively scant, and not much is known about tissue- or organism-specific glycosylation patterns.
In ASIC, the knuckle is largely uninvolved in gating and is considered a rather inert part of the palm/knuckle scaffold. In contrast, ENaC’s knuckle is an important source of heterogeneity between the three subunits. The carboxyl terminus of α6 differs between the three subunits, and its interface with the adjacent subunit’s finger domain may contribute to the enforcement of a clockwise subunit arrangement21.
2.3.2.2 Gating — the finger, thumb, and GRIP
The finger, thumb, and gating relief of inhibition by proteolysis (GRIP) domains form the essential regulatory machinery of the channel. The finger domain (blue) comprises three α-helices (α1–3). The thumb domain (green) comprises two large helices, α4 and 5.
The α- and γGRIP domains (cyan) are, as the name painstakingly makes clear, expected to block channel opening until they are proteolytically excised (see Section 2.4.1). The βGRIP is not cleaved during canonical channel opening and is, perhaps relatedly, the GRIP for which we have the best structural information. We can model a sheet of two anti-parallel pairs of β-strands in βGRIP. The GRIP domain shrouds the interface between the finger and thumb, forming extensive contacts between the two.
In ASIC, protonation of an acidic pocket formed by the β-ball, finger, and thumb of a single subunit causes collapse of the thumb toward the finger. This pulls the palm (and therefore the TMD) away from the symmetry axis, opening the channel. In ENaC, no such acidic residues exist. Instead, an aromatic pocket is formed by the finger, thumb, and GRIP domains. Based on this similar architecture and a slew of cross linking studies, it is thought that the GRIP domain forms a wedge between the finger and thumb, preventing their collapse31–33.
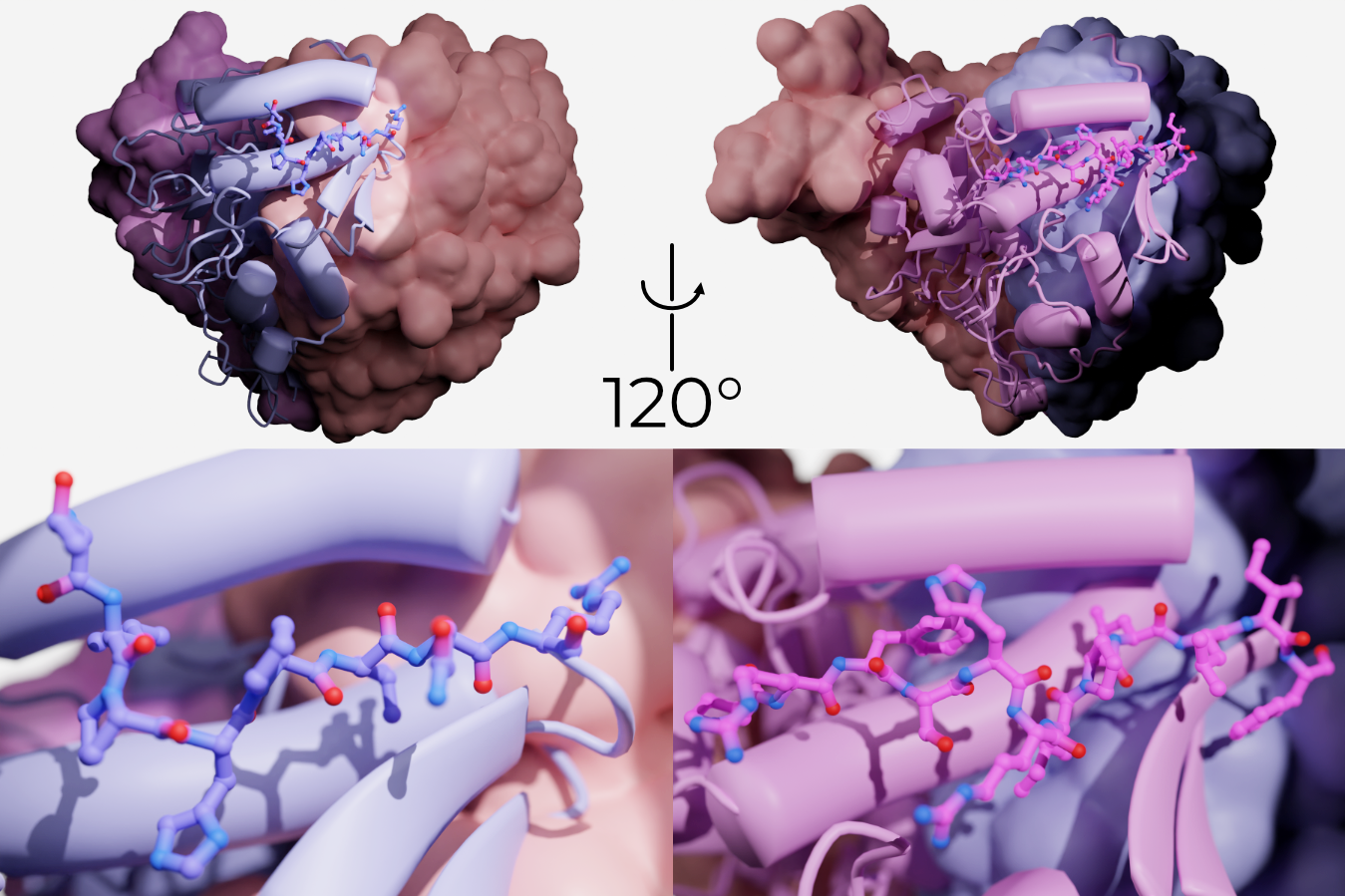
Indeed, the Kleyman laboratory has identified short segments of the GRIP domain (eight and eleven amino acids in α and γENaC, respectively) which, when flowed over open channels, close them (Figure 2.9)34–36. These peptides are part of the P1 strand of the GRIP domain and pack tightly against the remaining GRIP domain, α2 of the finger domain, and the upper edge of the thumb.
We have hypothesized that cleavage and subsequent removal of the P3 strand releases P1 and, therefore, decouples the finger and thumb21. From here, we and others have assumed that the gating mechanism is more-or-less that of ASIC: the finger and thumb collapse, and their movement pulls the palm and TMD away from the pore axis (Figure 2.3). However, without a map and model of the open and closed channels, we cannot rule out other mechanisms.
The new results from FaNaC call this assumption into question26. The finger and thumb do not collapse in FaNaC. Rather, they move in a concerted action up and away from the membrane, toward the α6 helix of the adjacent knuckle domain.
It is worth noting here that while FaNaC is gated by peptide binding (as is ENaC, after a fashion), FaNaC’s ligand binding pocket is distinct from ENaC’s. The FaNaC peptide binds a pocket comprising α2, the β6-β7 linker, and α6 from the adjacent subunit, i.e., the finger and knuckle. ENaC’s GRIP domain interacts instead with the thumb and finger. Since the FaNaC pocket is on “the other side” of the finger, it would not be entirely surprising if the movement of the finger in the two channels is different.
A histidine in the β9-α4 loop of FaNaC (which connects the palm and thumb) points down between TM1 and TM2. Mutation of this histidine to serine results in constitutively active channels, implying that the energy gained by ligand binding is used to overcome the interaction of this histidine with the TMs. ASIC and ENaC both have bulky residues (tryptophan and tyrosine, respectively) in this position.
2.3.2.3 Conduction — what we expect for the TMD
No map of ENaC has reliable density for the transmembrane domain. Nevertheless, we can infer a great deal from electrophysiological studies of ENaC and maps of other members of the ENaC/DEG family, mostly the well-studied ASIC.
ASIC’s gating cycle comprises three states: resting, open, and desensitized (Figure 2.1). The pore of the first solved structure of ASIC was asymmetric, but this truncated crystallization construct did not conduct sodium currents37. Subsequent structures of ASIC show symmetric pores throughout its gating cycle.
In the resting state of ASIC, negative charges clustered in the acidic pocket (formed by the finger, thumb, and β-ball) hold the finger and thumb apart. The separation of the thumb from the finger pushes the palm inward, holding the channel in a position which obstructs its lateral gate.
When ASIC encounters low extracellular pH, the residues in the acidic pocket are protonated. This relieves electrostatic repulsion and allows the finger/thumb interface to collapse, pulling the palm and TMD inward and opening a lateral fenestration just above the membrane (Figure 2.4)7.
Mutating a residue just above the second transmembrane helix in βENaC holds the channel open, likely by holding open a similar fenestration38. This βS520K is known as the DEG mutation, named after the C. elegans channel from which the mutation is derived.
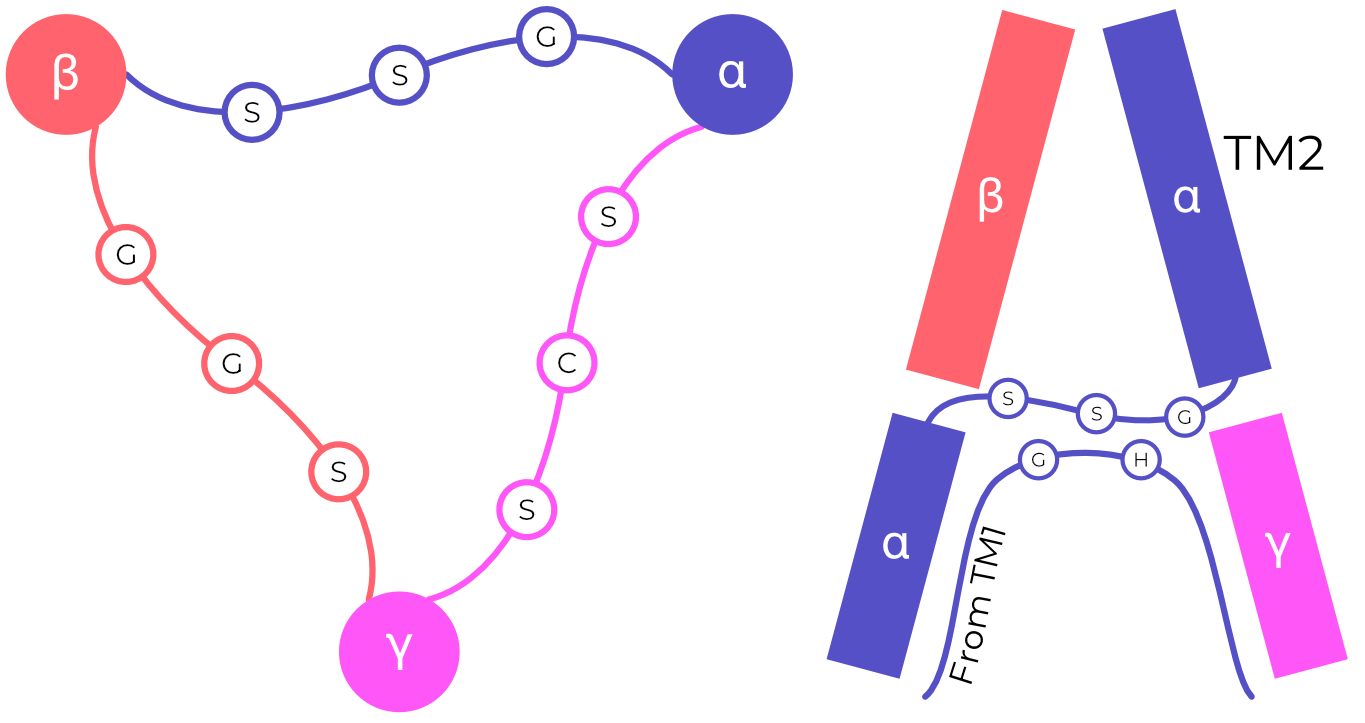
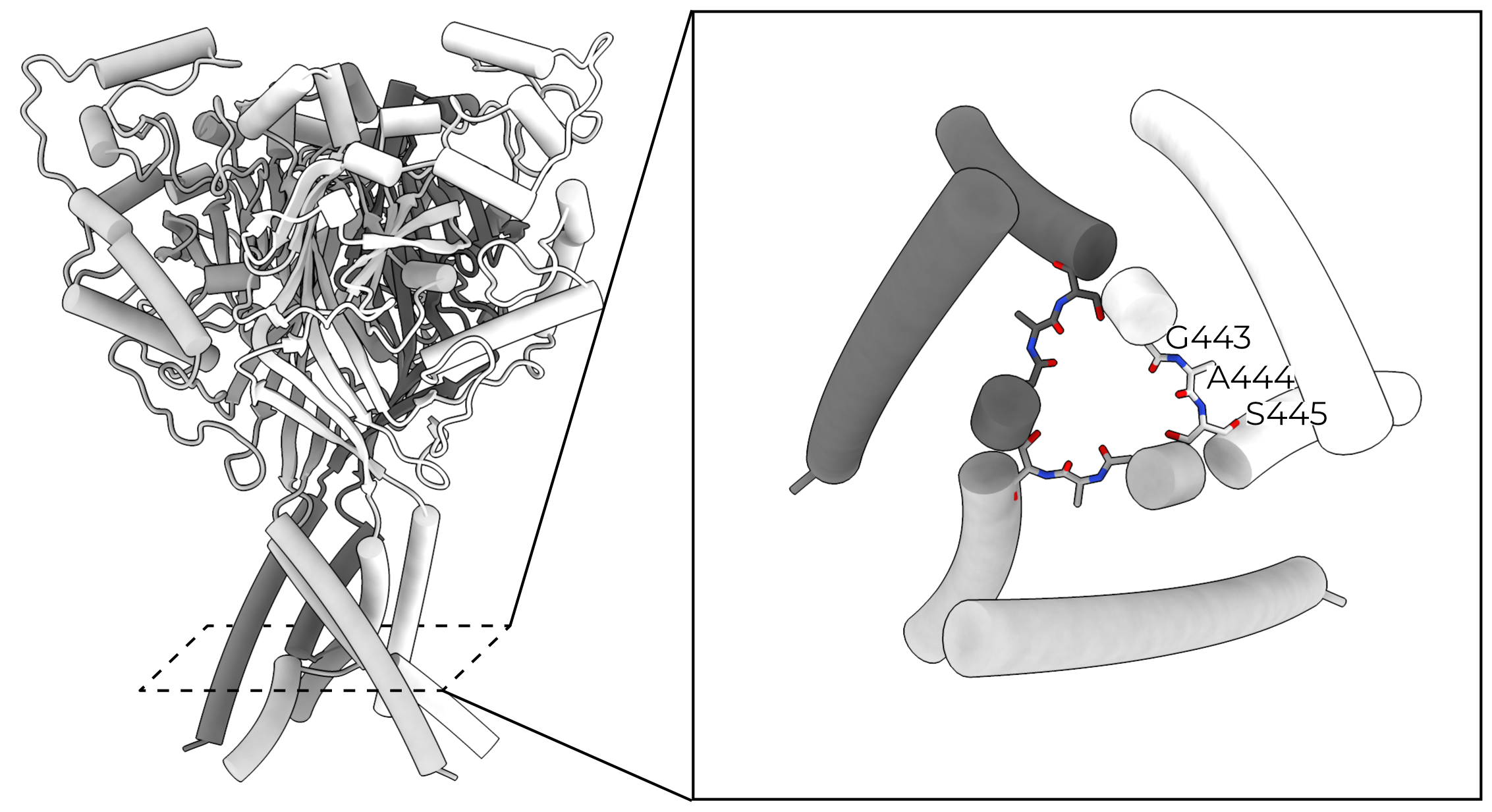
Interestingly, TM2 is split halfway through the membrane, with the C-terminal side swapping to complete the TM2 of the adjacent (clockwise) subunit. This domain swap forms a triangular arrangement of short Gly-Ala-Ser loops, called the GAS belt. The GAS belt forms ASIC’s selectivity filter40. The homologous residues in the α-, β-, and γENaC (GSS, GGS, and SCS respectively, Figure 2.10) play a significant role in that channel’s selectivity, implying the same pore structure and domain swap4,6,41–43.
In the open state, the ions pass from the extracellular fenestration down a negatively-charged pore through the GAS belt, and into the intracellular fenestration. The GAS belt selects for Na+ over other cations by a “sieving mechanism”, physically preventing the passage of ions with hydrated radii too large for the filter. It is worth noting here that ENaC’s significantly higher Na+ selectivity and lower unitary currents suggest that, unlike ASIC, Na+ ions are dehydrated before passing through ENaC44. We thus expect that ENaC’s selectivity filter will be significantly narrower than that of ASIC45.
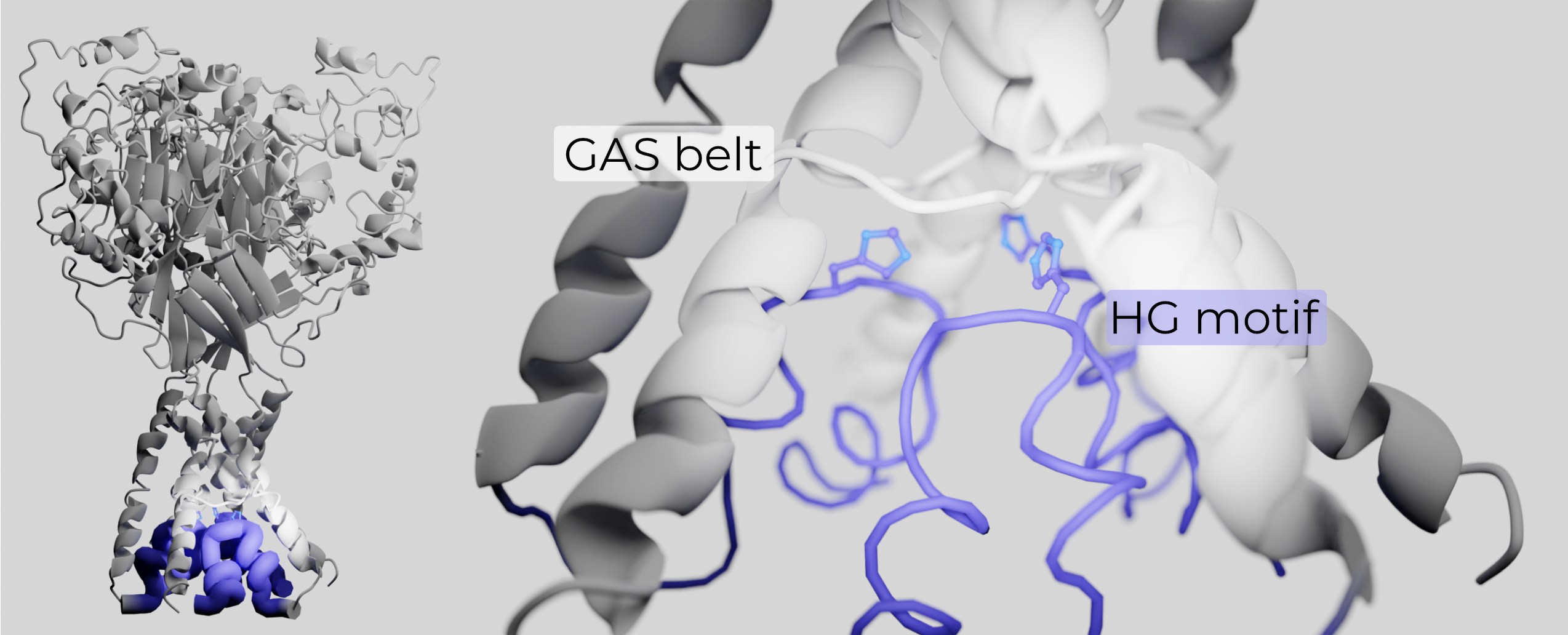
The conical region below the GAS belt and between TMs was believed to be filled with unstructured lipids. However, a recent structure of ASIC revealed that the N-terminus re-enters the membrane to buttress the GAS belt (Figure 2.11)46. The re-entrant loop also houses the highly conserved N-terminal two-residue His-Gly pair (the HG motif). The HG motif is critically important in gating and selectivity of ENaC, and likely plays a similar role in other ENaC/DEG family members47–49. The HG motif supports the GAS belt through several hydrogen bonds and hydrophobic interactions with adjacent TM2s, stabilizing this otherwise-fragile structure.
Unlike ENaC, ASIC desensitizes. In desensitized ASICs, the β11-β12 linker in the palm domain is flipped toward the channel center29. This decouples the still-collapsed finger and thumb domains from the lower palm and TMD, allowing the fenestration to close (Figure 2.1). In essence, the desensitized channels have an open ECD but a closed TMD. As ENaCs do not desensitize, the role of this linker is likely different. However, the linker sequence is highly conserved between ASIC and ENaCs, and an L→Q mutation results in a significant increase in current50.
2.3.3 Glycosylation
Glycans play an essential role in channel trafficking and maturation. The β subunit is heavily glycosylated, with eleven predicted sites in human compared to α and γ’s six and five, respectively. Prior to this work, four sites on βENaC, two on αENaC, and one on γENaC had been modeled21. Deletion of all glycosylation sites on any of the three subunits significantly decreases both total and surface expression of ENaC51. The same dependence of expression on glycosylation is seen in ASIC52.
ENaC glycosylation has functional implications beyond surface expression. Glycan maturation and proteolytic processing appear to happen in a concerted fashion, but they are not interdependent53. SSI is significantly reduced in ENaCs without glycans on any of the three subunits51. ASIC also depends on glycans for proper channel function — deletion of specific glycans changes current magnitude or proton affinity52.
To my knowledge, investigation of ENaC glycosylation has thus far been limited to knock-out of all predicted glycosylation sites in a single subunit or the entire channel. A study of single-glycan knock-outs, as has been performed in ASIC, would be invaluable in determining the specific contribution of glycosylation of each ECD domain.
2.4 Direct regulation of ENaC
2.4.1 Proteases
Proteolytic activation of ENaC was discovered shortly after the channel itself. Overnight treatment of Xenopus laevis bladder with aprotinin (a serine protease inhibitor) dramatically reduced ENaC currents; the same effect was observed in an X. laevis cell line54,55. ENaC currents were quickly rescued by treatment with the serine protease trypsin. Thus, an activating role for proteases was established.
ENaC exists in two populations at the cell surface: “near-silent” channels with a low PO (less than 0.02) and activated channels which are essentially constitutively open (PO ≅ 0.9)56,57. Treatment with exogenous trypsin activates the near-silent channels, and cleaved and uncleaved ENaC is found in cells and extracellular vesicles (making a secondary messenger unlikely)58–61.
Prostasin was the first protease found to activate ENaC55. Furin’s role in channel activation was reported shortly thereafter59. Prostasin is a membrane-associate serine protease, expressed highly in prostate and seminal fluid and moderately in other tissues including colon, lung, and kidney62. In contrast to prostasin’s relatively circumscribed expression, furin (also a serine protease) is expressed in essentially all human tissues. Furin plays a role in a vast array of biological processes including ENaC activation and is well-reviewed by Thomas63.
Both furin and prostasin are enteropeptidases which cleave following polybasic tracts (i.e., stretches of amino acids with multiple Lys or Arg residues). Both αENaC and γENaC have canonical cleavage sites in their GRIP domains which open the channel (Section 2.3.2.2)59. Current research suggests that, during channel activation, furin cuts α twice and γ once, while prostasin cuts γ once.
Unfortunately, these sites are in regions we have not yet been able to resolve. The relative changes in PO resulting from each cleavage event is still unclear, and seems to be tissue dependent12,59,64. In addition to prostasin, γENaC is also cleaved and activated by a wide repertoire of different proteases in different tissues64–74.
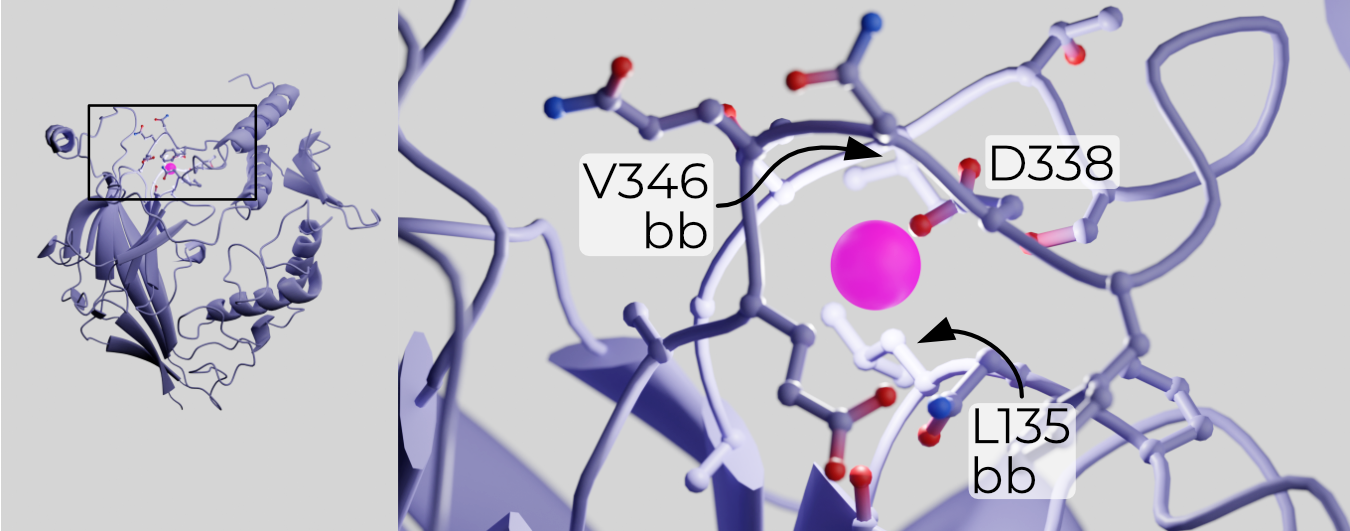
Removal of the inhibitory peptides is thought to open ENaC by relieving SSI19. Mutagenesis and structural work identified a putative Na+-binding “acidic pocket” formed by the finger and β-ball (Figure 2.12)20,21. If the GRIP domain does indeed prevent collapse of the finger and thumb toward each other, Na+ binding to a low-affinity site on the other side of α1 may pull the finger up and away from the thumb. Structures demonstrating such movement have yet to be captured — indeed, all structures solved before this work were in high-Na+ buffers.
Proteolytic activation has been described in great detail in a wide range of in vitro systems, but the link is not quite so obvious in vivo. For instance, mice treated with aprotinin show reduced ENaC activity as expected75. However, when mice are fed a low-sodium diet, the appearance of cleaved ENaC and increases in amiloride-sensitive current are largely independent76. It is thus not obvious that the direct cleavage/current relationship observed in vitro is true in vivo. What’s more, while proteolytic cleavage may be the main pathway by which ENaC is opened, ENaC is also sensitive to other signals.
2.4.2 The membrane
2.4.2.1 Phospholipids
Like many ion channels, ENaC is modulated by phospholipids. For instance, ENaC is known to segregate into lipid rafts77,78. Cholesterol alone does not activate the channel, but it does facilitate the interaction between phosphatidylinositol and binding sites in the β and γ subunits80,81. This interaction is absolutely required for ENaC opening.
Treatment of inside-out patches with phosphatidylinositol bisphosphate (PIP2) increases ENaC PO within seconds, and PO is reduced again upon PIP2 removal, ruling out the possibility of PIP2-induced proteolytic cleavage82,83. Similarly, no change in channel number is observed in these experiments. A correlation between ENaC currents and PIP2 levels is also observed in mouse trachea84. Note that PIP2 only augments ENaC currents up to a point — more PIP2 than is necessary does not result in greater ENaC currents. Put another way, a lack of PIP2 reduces ENaC currents but PIP2 does not itself potentiate them85.
Unlike PIP2, phosphatidylinositol triphosphate (PIP3) does augment ENaC currents both directly and via a signaling cascade. In its immediate effect, PIP3 binds γENaC just after the second transmembrane domain to rapidly open the channel85–87. PIP3 also increases the number of channels at the cell surface in a longer cascade mediated by SGK (Section 1.2.3)88. In fact, aldosterone’s effect on SGK is mediated by PI3K and PIP389.
ENaC’s dependence on phosphatidylinositols presents a problem. PIPs are rare enough in the membrane that, if the lipids and channel were both freely diffusing, the expected PO of ENaC would be far lower than what is observed90,91. MARCKS is a lipid-binding protein which sequesters PIP2 and PIP3 into regions of high local concentration92. MARCKS also binds ENaC, potentially anchoring the channel in a region of the membrane with a high enough local concentration PIPs for the proper channel PO93.
2.4.2.2 Palmitoylation
Palmitoylation is a reversible process in which hydrophobic chains are attached to cysteine residues, anchoring them in the membrane. Multiple cysteines in β- and γENaC (but none in αENaC) are palmitoylated, with varying effects96–98.
Blocking palmitoylation of two cysteines in βENaC (in mouse: C43 and C557) significantly reduces ENaC currents96. This effect is not mediated by differences in trafficking or cleavage but by changes in channel PO, specifically increased SSI. C43 is suggestively close to the HG motif; it may be that membrane anchoring of this cysteine aids in proper folding of the re-entrant loop and therefore the pore.
γENaC also has two palmitoylated cysteines, C33 and C41, which straddle that subunit’s HG motif98. Again, C→A mutation of these cysteines increase SSI without affecting surface exposure or channel cleavage. Surprisingly, addition of a β palmitoylation mutant to a mutant γ background does not further reduce currents — put another way, the γ mutations are dominant.
2.4.3 Mechanosensation
Whether or not ENaC directly senses membrane stretch has long been a point of contention, but decades of careful research indicate that laminar sheer stress (LSS) opens the channel regardless of proteolytic state. Bath perfusion of X. laevis oocytes or CHO cells expressing ENaC produces amiloride-sensitive Na+ currents without changing their cleavage state99,100.
Single channel recordings indicate that LSS currents result from directly increased PO101. Although LSS can open uncleaved channels, pre-treatment of human ENaC with trypsin significantly primed LSS response101,102. Over the course of minutes, ENaC becomes less sensitive to repeated stimulation with LSS103. Mutant ENaCs with truncated C-termini did not show this response, indicating that it may be mediated by Nedd4-2 (Section 1.2.2.2).
Multiple domains may participate in LSS sensation. Genetic deletion of the GRIP domain slows response to LSS, although this may be an artifact of differences in finger/thumb coupling rather than a true measure of the GRIP domain’s participation in gating104. Work on human and rat ENaC demonstrated a strong dependence of LSS currents on glycosylation of the αENaC palm and knuckle; in another study, deletion of any single subunit’s glycans has no effect on LSS currents51,105. The former study makes a compelling argument that the glycans influence LSS by interacting with the extracellular matrix — a difference in oocyte preparation may therefore explain this discrepancy.
2.5 Conclusion
I’ve demonstrated that although we can make some assumptions about how changes in the ENaC ECD are coupled to pore opening, much of the channel’s gating mechanism remains unknown. For instance: which glycans are involved in which aspects of channel gating? How does sodium inhibit the channel, and where does it bind? Does ENaC contain a GAS belt and a lateral fenestration like ASIC?
Moreover, the existence of several, semi-overlapping means of modulating channel PO indicates that there may be multiple, physiologically relevant means of opening ENaC that cells use to fine-tune their Na+ permeability. At a more basic level, structures of ENaC’s TMD, pore, and selectivity filter would contribute to our understanding of the important ENaC/DEG family more broadly. It is with this in mind that I set out on the goals of this project: to solve the structure of the ENaC TMD, and to solve high-resolution structures of the channel in various salts.